Introduction
The fascinating process of facial nerve embryogenesis and its high susceptibility to a wide range of harmful factors, including viruses, bacterial infections, and genetic mutations, have strengthened our belief that even the most sophisticated imaging methods cannot provide a better understanding of facial nerve formation and functionality than the investigation of its development in human embryos.
The rhombencephalon develops from an excessively segmented neuroepithelial area of the cephalic portion of the neural tube, giving rise to neuromeres [1]. Müller and O’Rahilly proposed the first classification of the neuromeres, according to their position and appearance in the brain [2]. Thus, at Carnegie stage 9, the scientists identified 6 primary neuromeres, but by stage 14, all the secondary neuromeres had formed. The neuromeres were detectable until Carnegie stage 17. Lumsden revealed a distinct position of the cranial nerve roots in relation to individual rhombomeres in chick embryos. Thus, the trigeminal nerve root lies in r2, the facial nerve root in r4, and the glossopharyngeal in r6 [3]. The roots of the nerves were distinguished in the central furrow of the rhombomere, and "a tight correspondence between the position of each branchial motor nucleus and the rhombomeric pattern was observed" [3]. In vertebrates, the development of the face is related to the neural crest and is regulated by a "code of HOX genes in a segment-restricted way" [4]. By stage 17, the "branchial motor nerves emerge from nuclei within consecutive pairs of rhombomeres, each pair lying in register with an adjacent branchial arch" [5, p. 331], and a two-segment periodicity is established [5-7]. Under normal conditions, the trigeminal nerve develops from r2 and r3, the facial nerve from r4 and r5, the glossopharyngeal nerve from r6 and r7, and the vagus and accessory nerves from r7 and r8 [1-5, 8, 9].
In chick and mouse embryos, a particular HOX gene–GHOX 2.9 in chicks and HOX 2.9 in mice–is highly expressed at the level of r4 and the hyoid arch, from which the facial nerve derives [10-12]. The expression of HOX genes in the neural tube and neural crest cells, as well as the combined expression of HOX genes in both the neurogenic neural crest and branchial arches, determines the development of the facial nerve [9].
According to Sataloff and Danilo, in the third week of gestation, the facio-acoustic primordium derives from the rhombomeres of the rhombencephalon, giving rise to the facial nerve [13, 14]. The trunk of the facial nerve passes through the mesenchyme towards the facial neurogenic placodes, contributing to the formation of the nerve ganglia associated with the pharyngeal arches. By the fourth week of embryonic development, the facial nerve nucleus is formed, and the motor fibers of the facial nerve extend from it [13-15], while Müller considers that the motor nucleus of the facial nerve is not definitely formed until the last stage of embryogenesis (Carnegie stage 23) [2]. The parasympathetic ganglia derive from neural crest cells, and the axons of those neurons form the facial nerve [16]. In the 4th week of development, the chorda tympani is distinguished [13, 14]. During the 5th–7th weeks of gestation, the geniculate ganglion, the intermediate nerve, and the greater petrosal nerve appear [16, 17].
The geniculate ganglion is of mixed embryological origin, with the majority of its neurons deriving from the hyomandibular epibranchial placode, while the supporting cells are derivatives of the neural crest [18, 19]. In relation to the vestibulocochlear complex, the geniculate ganglion is located at the level of r4, being in contact with the epipharyngeal placode of the 2nd visceral arch [20].
Development of the motor components of the cranial nerves derived from the first three visceral arches depends on HOX gene expression [1]. Development of the facial nerve and of the geniculate ganglion "involves neuronal survival, differentiation, and neuron-glia interaction", regulated by the gene-related family of neurotrophins–nerve growth factor–which determines the growth of sympathetic and some sensory neurons, as well as neuronal survival [18]. During the 7th and 8th weeks, the mimic muscles derive from the second branchial arch, and by the 11th week of development, the facial nerve, which supplies these muscles, is characterized by obvious arborization. In the newborn, the anatomy of the facial nerve shows many similarities to that of the adult, except in the mastoid region, where the nerve is located more superficially due to underdevelopment of the mastoid process.
Considering the above-mentioned facts, the goal of our study was to highlight the morphological specific features of facial nerve development for a better understanding of its branching patterns, connections, and functionality in adults.
Material and methods
This current study was conducted on the basis of the Bilateral Agreement between the Department of Anatomy and Clinical Anatomy of Nicolae Testemițanu State University of Medicine and Pharmacy of the Republic of Moldova and the Department of Normal Anatomy of the Belarusian State Medical University in Minsk.
Fifty-two groups of serially sectioned human embryos and fetuses from the embryo-fetal historical collection of the Department of Normal Anatomy of the Belarusian State Medical University in Minsk were investigated. The crown-rump length (CRL) of the examined embryos and fetuses ranged from 4 mm to 70 mm.
According to available data, prior to dissection, the embryos and fetuses were fixed in 10% formalin solution, after which they were dissected by a microtome in sagittal, frontal, and transverse planes. The histological samples were subsequently stained using hematoxylin-eosin and Bielschowsky-Bücke silver impregnation methods. All the samples used in the current study were fixed and stained by the staff of the Department of Normal Anatomy of the Belarusian State Medical University in Minsk, at the time of the collection’s creation. The descriptive and morphometric parameters of the embryos and fetuses were stored in an Excel database, maintaining the coding of the original collection. The embryonic samples were classified according to Carnegie stages (day of gestation and CRL), as proposed by Robinson [21]. For description of the protocols, the OLYMPUS CX31 binocular microscope (eyepiece 10x, objectives 4x; 10x; 40x; 100x) and Nikon DS-Fi1 camera were used.
Results
At the end of Carnegie stage 13, a narrow furrow splits the facio-acoustic primordium into the facial and acoustic ganglia. At stage 15, the facial nerve exits the rhombencephalon as a straight, well-impregnated trunk, consisting of loosely arranged nerve fibers (Fig. 1A). The geniculate ganglion, chorda tympani, and greater petrosal nerves were also identified at stage 15.
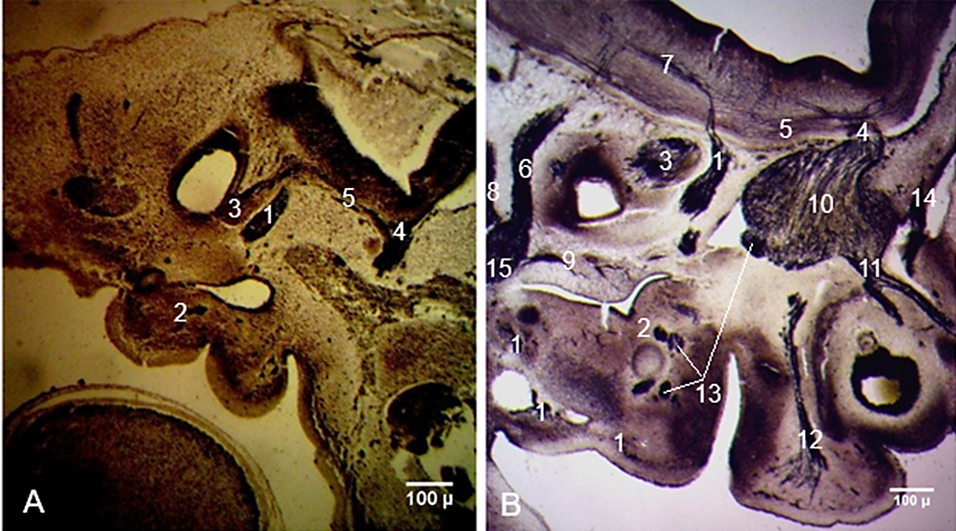 |
Fig. 1 Development and connections of the facial nerve in human embryos. A. Carnegie stage 15 (CRL 9 mm). B. Carnegie stage 19 (CRL 17 mm). Sagittal sections. Staining (Bielschowsky-Bücke and hematoxylin-eosin). Microphotographs. 1 – facial nerve; 2 – chorda tympani; 3 – acoustic ganglion (future vestibular and spiral ganglia); 4 – trigeminal nerve; 5 – intracerebral connections between the facial nerve and trigeminal nerve; 6 – glossopharyngeal nerve; 7 – intracerebral connections between the facial nerve and glossopharyngeal nerve; 8 – vagus nerve; 9 – tympanic nerve; 10 – trigeminal ganglion; 11 – ophthalmic nerve; 12 – maxillary nerve; 13 – mandibular nerve; 14 – trochlear nerve; 15 – inferior ganglion of the glossopharyngeal nerve. |
On deeper planes of the sagittal sections, the roots formed by the facial and trigeminal nerves faded, while those of the glossopharyngeal and vagus nerves were well distinguishable. The intracerebral connections between the facial and glossopharyngeal nerves were also identified. The connections between the vagus and hypoglossal nerves were highlighted. The neuronal groups that give rise to the facio-vestibulo-cochlear complex were detected in the region of the internal acoustic meatus at Carnegie stages 15-16 (CRL 9-11 mm).
At Carnegie stage 16, two roots of the facial nerve–an anterior and a posterior one–were highlighted on transverse cross-sections. The definitive formation of the intracerebral roots of the facial nerve and their fusion into a common trunk were established at Carnegie stage 18 (Fig. 2).
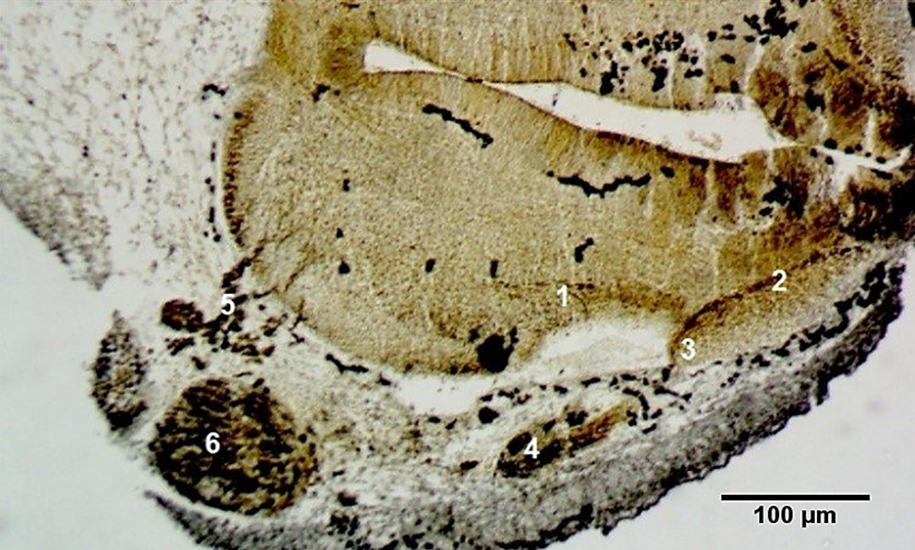 |
Fig. 2 Formation of the intracerebral roots of the facial nerve. Transverse cross-section through the hindbrain. Carnegie stage 18 (CRL 15 mm). Staining (Bielschowsky-Bücke). Microphotograph. 1 – anterior root of the facial nerve; 2 – posterior root of the facial nerve; 3 – facial nerve; 4 – geniculate ganglion; 5 – roots of the trigeminal nerve; 6 – trigeminal ganglion. |
At Carnegie stage 19, the intracerebral connections of the facial nerve with the trigeminal and glossopharyngeal nerves became thicker, and a clear fibrillar character of these connections was observed. The peripheral branches of the facial nerve were better distinguished, and their connections with the peripheral branches of the trigeminal and glossopharyngeal nerves became more obvious. The course of the tympanic nerve through the mesenchyme of the developing petrous part of the temporal bone was clearly identified. The trochlear nerve and all the primary branches of the trigeminal nerve were well distinguished (Fig. 1B).
The biotaxis of the neuroblasts from the depth of the rhombencephalon towards the external surface of the brain appeared as fibrocellular cords at Carnegie stage 16 (Fig. 3A). At the cranial end of the hindbrain, the roots of the trigeminal nerve were highlighted. Medial to the trigeminal nerve, the intracerebral path of the facial nerve was identified. The internal knee of the facial nerve was observed on frontal sections of embryos at Carnegie stage 23, which is the last stage of embryonic development (Fig. 3B). The fibers of the facial nerve entered the internal acoustic meatus, close to the vestibular ganglion, continuing their course within the facial canal, and, upon exiting the stylomastoid foramen, the nerve appeared as a thick fibrous cord.
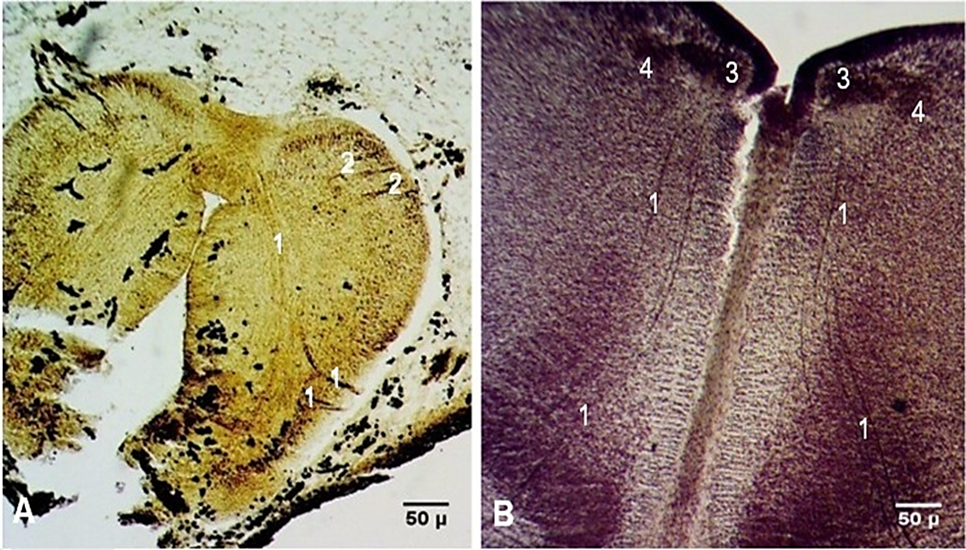 |
Fig. 3 Intracerebral course of the facial nerve in human embryos. A. Transverse cross-section. Carnegie stage 16 (CRL 11 mm). B. Frontal section. Carnegie stage 23 (CRL 27 mm). Staining (Bielschowsky-Bücke and hematoxylin-eosin). Microphotographs. 1 – fibers of the facial nerve; 2 – fibers of the trigeminal nerve; 3 – motor nucleus of the facial nerve; 4 – internal knee of the facial nerve. |
Peripheral connections of the facial nerve were distinguished starting from Carnegie stage 15 (CRL 9 mm). Among the earliest revealed connections were those of the chorda tympani with the lingual branches of the glossopharyngeal nerve (Fig. 4A). At Carnegie stage 16 (CRL 11 mm), a communicating branch of the facial nerve with the tympanic plexus was highlighted (Fig. 4B). The first appearance of the parotid plexus was observed at Carnegie stage 17 (CRL 14 mm), and multiple thin intraplexal connections between the main divisions of the future parotid plexus were distinguished (Fig. 4C). At Carnegie stage 21 (CRL 23 mm), the extracranial branches of the facial nerve were represented by multiple distal arborizations. A plexiform structure, formed by connections between the distal ramifications of the temporofacial division of the facial nerve and the infraorbital branch of the maxillary nerve, was marked out in the infraorbital region. The infraorbital plexiform structure consisted of a well-defined fibrillar component with highly complex connections, known as the pes anserinus minor (Fig. 4 D).
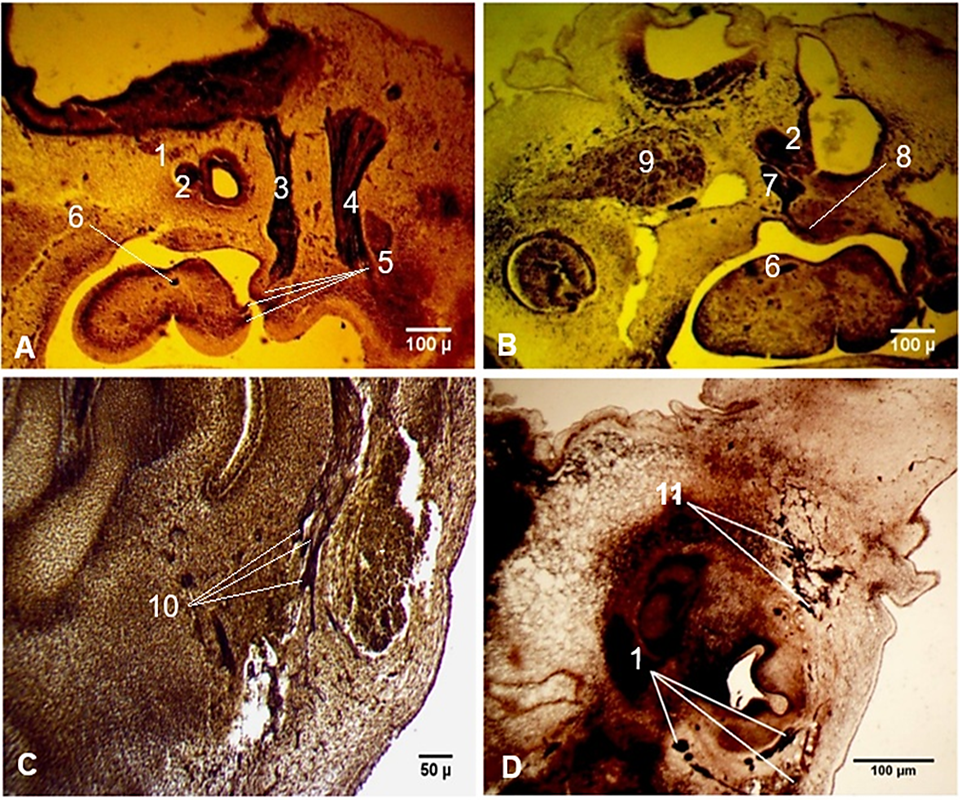 |
Fig. 4 Peculiarities of the peripheral connections of the facial nerve. A. Connections of the facial nerve with the lingual branches of the glossopharyngeal nerve. Carnegie stage 15 (CRL 9 mm). B. Communicating branch of the facial nerve with the tympanic plexus. Carnegie stage 16 (CRL 11 mm). C. Intraplexal connections between the branches of the parotid plexus. Carnegie stage 17 (CRL 14 mm). D. A plexiform structure in the infraorbital region. Carnegie stage 21 (CRL 23 mm). Sagittal sections. Staining (Bielschowsky-Bücke). Microphotographs. 1 – facial nerve; 2 – acoustic ganglion; 3 – glossopharyngeal nerve; 4 – vagus nerve; 5 – connections of the facial nerve with the lingual branches of the glossopharyngeal nerve; 6 – chorda tympani; 7 – geniculate ganglion; 8 – communicating branch of the facial nerve with the tympanic plexus; 9 – trigeminal ganglion; 10 – intraplexal connections between the branches of the parotid plexus; 11 – plexiform structure in the infraorbital region, formed by connections between the facial and trigeminal nerves (pes anserinus minor). |
As a specific feature of the geniculate ganglion during its embryonic and fetal development, continuous modifications in its shape–from oval to triangular and ovoid-triangular–were noted. A series of microscopic morphological changes were also observed. At Carnegie stage 18, the geniculate ganglion had an ovoid shape and was predominantly formed by nerve fibers, among which growing neuroblasts were distinguished. An hourglass-shaped narrowing was revealed on the medial side of the ganglion. The neuroblasts were randomly distributed within the geniculate ganglion; nevertheless, their concentration was clearly higher along its margins and in the core of the ganglion (Fig. 5A).
In fetuses with a CRL of 55 mm, the geniculate ganglion was oval, and the hourglass-shaped narrowing had disappeared. A neuronal redistribution within the geniculate ganglion occurred: the neuroblasts surrounded the external margin of the ganglion as a thick collar, while the central part consisted predominantly of nerve fibers, with only a few thin cords of neuroblasts highlighted in its core (Fig. 5B).
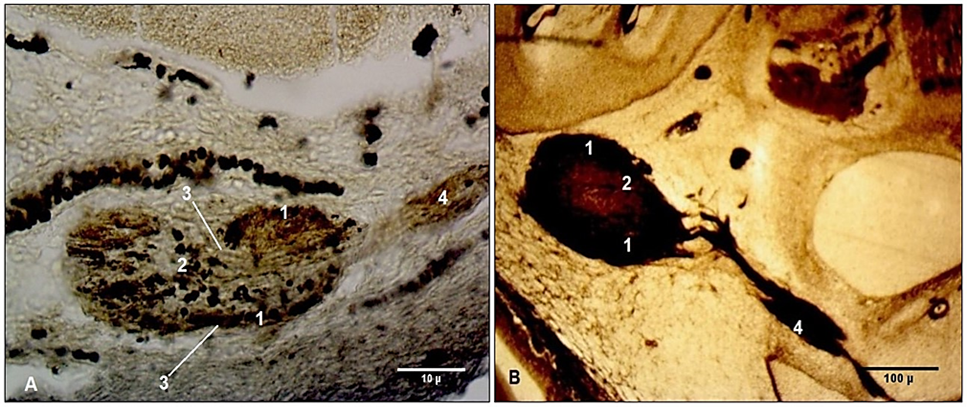 |
Fig. 5 Developmental peculiarities of the geniculate ganglion in embryos and fetuses. Transverse cross-sections. A. Embryo at Carnegie stage 18 (CRL 15 mm). B. Fetus (CRL 55 mm). Staining (Bielschowsky-Bücke). Microphotographs. 1 – neuroblasts located at the periphery of the geniculate ganglion; 2 (A) – central cluster of neuroblasts; 2 (B) – thin cords of neuroblasts in the core of the ganglion; 3 – hourglass-shaped narrowing of the ganglion; 4 – facial nerve. |
Among the specific features of the geniculate ganglion were its connections with the vestibular ganglion. At Carnegie stage 20, a thin connection between the geniculate and vestibular ganglia was highlighted. In fetuses, those connections became thicker and stronger (Fig 6).
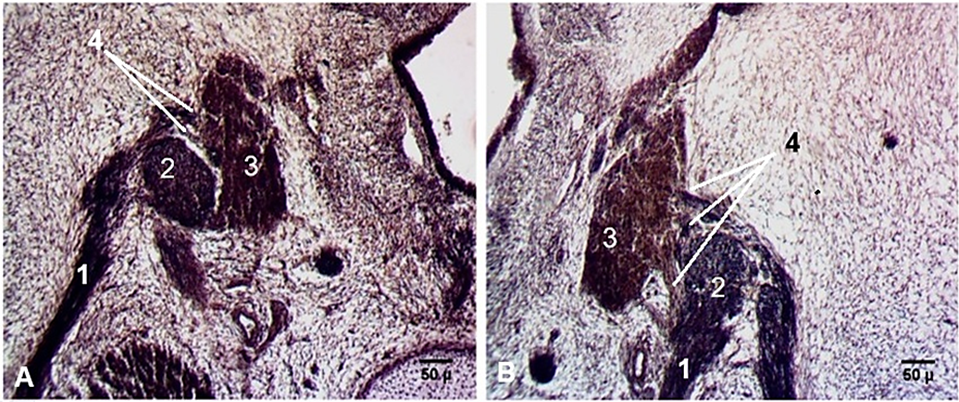 |
Fig. 6 Connections between the geniculate and vestibular ganglia. Transverse cross-sections. Fetus (CRL 57 mm). A. Left side. B. Right side. Staining (hematoxylin-eosin). Microphotographs. 1 – facial nerve; 2 – geniculate ganglion; 3 – vestibular ganglion; 4 – connections between the geniculate and vestibular ganglia. |
Two abnormalities of the facial nerve were found among the examined embryos and fetuses. On a sagittal section of an embryo at Carnegie stage 19 (CRL 17 mm), a double, overlapping enlargement of the geniculate ganglion, from which two separate facial trunks derived, was detected (Fig. 7A). Each facial trunk distally divided into multiple branches, giving rise to the parotid plexus. The peripheral branches of the anterior trunk were directed anteriorly, while those of the posterior trunk were spread both anteriorly and posteriorly, and a connection between these two trunks was revealed. In a fetus (CRL 57 mm), an aberrant branch to the sternocleidomastoid muscle, considered casuistic, was found (Fig. 7B).
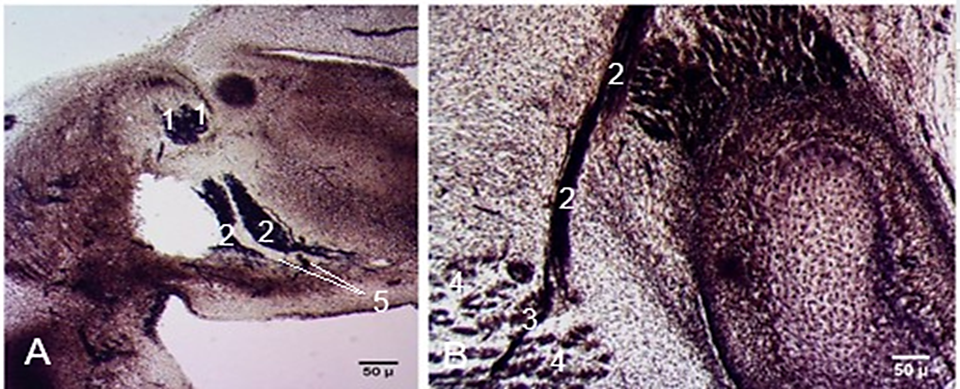 |
Fig. 7 Abnormal development of the facial nerve. A. Double facial nerve trunk. Sagittal section. Embryo at Carnegie stage 19 (CRL 17 mm). B. Aberrant branch from the facial nerve to the sternocleidomastoid muscle. Transverse cross-section. Fetus (CRL 57 mm). Staining (hematoxylin-eosin). Microphotographs. 1 – geniculate ganglion; 2 – facial nerve; 3 – aberrant branch to the sternocleidomastoid muscle; 4 – sternocleidomastoid muscle; 5 – connections between the facial nerve trunks. |
Discussion
In the third week of embryonic development, no branches of the facial nerve or ganglia are distinguished; however, the facio-acoustic primordium is already formed [13, 14, 22].
At Carnegie stage 13, the facial nerve forms "a complex structure with the vestibulocochlear ganglion", which, by stage 14, divides into separate ganglia [20]. In the current study, the facio-acoustic primordium in the third week of embryonic life appeared as a triangular structure, separated by a narrow furrow into the facial nerve and acoustic ganglion.
A "fan-shaped pattern" of the facial nerve exit from the hindbrain nuclei located at the level of r4 and r5 was described in mice [23]. The fibers exiting the brain from the nuclei at the level of r4 entirely contributed to the formation of the facial nerve trunk. Two groups of nuclei were identified at the level of r5–a medial and a lateral one–and only the axons of the medial nuclear group participated in the formation of the motor root of the facial nerve. The axons of the lateral group of nuclei were considered to be preganglionic parasympathetic fibers [23].
Bruska et al. noted that, at stage 13, the geniculate ganglion consisted of vertical rows of cells running parallel to the facial nerve fibers, and the ganglion appeared as a separate morphological structure [24, 25]. According to Gasser, "the geniculate ganglion is well defined in 11.0-13.5 mm embryos" [26]. In our series of embryos, the geniculate ganglion appeared as a separate structure in embryos with a CRL of 11 mm. A specific feature of the geniculate ganglion was its tendency to change shape–from ovoid to fusiform and triangular–although the oval shape was generally characteristic of the ganglion. In the investigated samples, the central part of the geniculate ganglion consisted of nerve fibers, laterally bounded by two nerve cords of neuroblasts. In the core of the ganglion, parallel rows of neuroblasts were identified. At stage 18, the neuroblasts were large, and many more nerve fibers were present in the geniculate ganglion compared with the acoustic ganglion.
Danilo emphasized that the external knee of the facial nerve begins its formation during the sixth week of development [14]. In the current study, changes in the course of the facial nerve and its bending around the otic capsule were observed during the same period of gestation. The facial nerve emerged from the brain as many thin fibers that continued into the facial nerve trunk, from which the chorda tympani nerve at stage 14 and the greater petrosal nerve at stage 15 originated. Similar results were reported by Lobko [27].
According to Gasser, the peripheral divisions of the facial nerve began the parotid plexus formation in 18 mm embryos, while in our investigations the secondary divisions of the facial nerve were observed in embryos with a CRL of 14 mm, and some of those branches spread distally in a network fashion [26].
It is considered that migration of the pharyngeal efferent neurons influences the formation of the facial nerve internal knee, which appears at Carnegie stage 23 [25]. In our study, the internal knee of the facial nerve was also observed at Carnegie stage 23.
Experimental studies have proved that the zinc finger gene Krox-20 is a segment-specific regulator of HOX gene expression [9]. A similar regulatory effect on HOX gene expression is exerted by retinoic acid [28]. Under the influence of retinoic acid and the zinc finger Krox-20 gene, a transformation of the motor portion of the trigeminal nerve into the facial nerve was observed. The r2/3 transformed into r4/5, and subsequently the entire portion of the trigeminal area transformed into the facial-vestibuloacoustic area. Changes in HOXB1 gene expression resulted in duplications of r2/3 into r4/5, but no changes were noted in the normal r4/5 segments [9, 28].
Neural crest cells are multipotent and highly migratory. They are influenced by a range of cell signaling factors such as Fibrobrlast Growth Factor, Bone Morphogenetic Proteins, retinoic acid, Notch ligands, Receptor Tyrosine Kinases, and Wnt proteins. These signaling pathways are active in the neural placodes and neural cells precursors that give rise to the cranial nerves. Disturbances in neural cell migration and differentiation can lead to various malformations and aberrant neurogenesis of the cranial nerves [29], including variations in the number and course of the facial nerve trunk [30]. The aberrant branch of the facial nerve to the sternocleidomastoid muscle could result from fusion of the posterior belly of the digastric muscle with the sternocleidomastoid muscle [31].
Conclusions
The development of the facial nerve is influenced by a range of factors, among which are HOX genes, neural crest cell migration and differentiation, signaling molecules, and viral and bacterial infections during the first 8 weeks of pregnancy. Early specific features of facial nerve development include: 1) intracerebral and peripheral connections with the neighboring cranial nerves; 2) intraplexal connections of the parotid plexus branches; 3) distinguishable intracerebral pathways; 4) connections of the geniculate ganglion with the vestibular ganglion. As late developmental peculiarities, the following were identified: 1) changeable macro- and microstructure of the geniculate ganglion; 2) appearance of the pes anserinus minor; 3) formation of the motor nucleus, internal knee, and intracerebral pathways. The developmental specific features of the facial nerve are obvious evidence of its complex morphology in adults.
Competing interests
None declared.
Authors’ contributions
AB – conception and design of the work. AB, NT – contribution to acquisition of data. AB, ZZ, IC, NT – contribution to analysis and interpretation of data. AB, ZZ, IC, NT, ABend, NO, SL – drafting the article. All authors reviewed the work and approved the final version of the manuscript.
Acknowledgements and funding
No external funding.
Ethics approval
This research project was approved by the Research Ethics Committee of Nicolae Testemițanu State University of Medicine and Pharmacy of the Republic of Moldova (Minutes no. 1 of 19.09.2014) and complies with the 1964 Helsinki Declaration and its later amendments.
Provenance and peer review
Not commissioned, externally peer-reviewed.
Authors’ ORCID IDs
Angela Babuci – https://orcid.org/0000-0003-0305-1279
Zinovia Zorina – https://orcid.org/0000-0002-0749-6083
Ilia Catereniuc – https://orcid.org/0000-0002-5479-4198
Nataliya Trushel – https://orcid.org/0000-0003-0865-2495
Anastasia Bendelic – https://orcid.org/0000-0002-2838-3168
Nadia Ostahi – https://orcid.org/0009-0003-7197-0178
Sofia Lehtman – https://orcid.org/0000-0003-4653-8589
References
Trainor PA, Krumlauf R. Patterning the cranial neural crest: hindbrain segmentation and Hox gene plasticity. Nat Rev Neurosci. 2000;1(2):116-124. doi: 10.1038/35039056.
Müller F, O'Rahilly R. The timing and sequence of appearance of neuromeres and their derivatives in staged human embryos. Acta Anatomica. 1997;158(2):83-99. doi: 10.1159/000147917.
Lumsden A, Keynes R. Segmental patterns of neuronal development in the chick hindbrain. Nature. 1989;337(6206):424-428. doi: 10.1038/337424a0.
Hunt P, Wilkinson D, Krumlauf R. Patterning the vertebrate head: murine Hox 2 genes mark distinct subpopulations of premigratory and migrating cranial neural crest. Development. 1991;112(1):43-50. doi: 10.1242/dev.112.1.43.
Lumsden A. The cellular basis of segmentation in the developing hindbrain. Trends Neurosci. 1990;13(8):329-335. doi: 10.1016/0166-2236(90)90144-Y.
Kuratani SC, Eichele G. Rhombomere transplantation repatterns the segmental organization of cranial nerves and reveals cell-autonomous expression of a homeodomain protein. Development. 1993;117(1):105-117. doi: 10.1242/dev.117.1.105.
Carpenter EM, Goddard JM, Chisaka O, Manley NR, Capecchi MR. Loss of Hox-A1 (Hox-1.6) function results in the reorganization of the murine hindbrain. Development. 1993;118(4):1063-1075. doi: 10.1242/dev.118.4.1063.
Clarke JD, Lumsden A. Segmental repetition of neuronal phenotype sets in the chick embryo hindbrain. Development. 1993;118(1):151-162. doi: 10.1242/dev.118.1.151.
Wilkinson DG. Molecular mechanisms of segmental patterning in the vertebrate hindbrain and neural crest. Bioessay. 1993;15(8):499-505. doi: 10.1002/bies.950150802.
Frohman MA, Boyle M, Martin GR. Isolation of the mouse Hox-2.9 gene; analysis of embryonic expression suggests that positional information along the anterior-posterior axis is specified by mesoderm. Development. 1990;110(2):589-607. doi: 10.1242/dev.110.2.589.
Maden M, Hunt P, Eriksson U, Kuroiwa A, Krumlauf R, Summerbell D. Retinoic acid-binding protein, rhombomeres and the neural crest. Development. 1991;111(1):35-43. doi: 10.1242/dev.111.1.35.
Guthrie S, Muchamore I, Kuroiwa A, Marshall H, Krumlauf R, Lumsden A. Neuroectodermal autonomy of Hox-2.9 expression revealed by rhombomere transpositions. Nature. 1992;356(6365):157-159. doi: 10.1038/356157a0.
Sataloff RT. Embryology of the facial nerve and its clinical applications. Laryngoscope. 1990;100(9):969-84. doi: 10.1288/00005537-199009000-00011.
Danilo AGO. Facial nerve: embryology and anatomy of its nucleus. MOJ Anat Physiol. 2018;5(3):164-166. doi: 10.15406/mojap.2018.05.00183.
Babuci A, Catereniuc I, Zorina Z, Botnari T, Lehtman S, Nastas L. Peculiarities of the facial nerve development. In: Trushel’ N, editor. Innovatsii i aktual’nye problemy morfologii: Sbornik nauchnykh statei, posviashchennye 100-letiiu kafedry normal’noi anatomii Belorusskogo gosudarstvennogo meditsinskogo universiteta [Innovations and current problems of morphology: Collection of scientific articles dedicated to the 100th anniversary of the Department of Normal Anatomy of the Belarusian State Medical University], Oct 2021. Minsk: BGMU; 2021, p. 361-365.
Sadler TW. Langman's medical embryology. 12th ed. Philadelphia: Lippincott William &Wilkins; 2012; 384 p.
Babuci A. Development of the facial nerve in human embryos. In: MedEspera: the 8th International Medical Congress for Students and Young Doctors; 2020 Sep 24-26; Chisinau, Republic of Moldova: Abstract book. Chișinău; 2020. p. 231-232.
D'Amico-Martel A, Noden DM. Contributions of placodal and neural crest cells to avian cranial peripheral ganglia. Am J Anat. 1983;166(4):445-468. doi: 10.1002/aja.1001660406.
Vazquez E, Calzada B, Naves J, Garnacho SS, del Valle M, Vega JA, Represa J. Developmental changes in nerve growth factor (NGF) binding and NGF receptor proteins trkA and p75 in the facial nerve. Anat Embryol (Berl). 1994;190(1):73-85. doi: 10.1007/BF00185848.
Weglowski M, Wozniak W, Piotrowski A, Bruska M, Weglowska J, Sobanski J, Grzymislawska M, Lupicka J. Early development of the facial nerve in human embryos at stages 13-15. Folia Morphol. 2015;74(2):252-257. doi: 10.5603/FM.2015.0039.
Robinson HP. Sonar measurement of fetal crown-rump length as means of assessing maturity in first trimester of pregnancy. Br Med J. 1973;4(5883):28-31. doi: 10.1136/bmj.4.5883.28.
Sataloff RT, Selber JC. Phylogeny and embryology of the facial nerve and related structures. Part I: Phylogeny. Ear Nose Throat J. 2003;82(9):704, 707-710.
Goddard JM, Rossel M, Manley NR, Capecchi MR. Mice with targeted disruption of Hoxb-1 fail to form the motor nucleus of the VIIth nerve. Development. 1996;122(10):3217-3228. doi: 10.1242/dev.122.10.3217.
Bruska M, Ulatowska-Błaszyk K, Węgłowski M, Woźniak W, Piotrowski A. Differentiation of the facial-vestibulocochlear ganglionic complex in human embryos of developmental stages 13-15. Folia Morphol (Warsz). 2009;68(3):167-173.
Müller F, O'Rahilly R. The initial appearance of the cranial nerves and related neuronal migration in staged human embryos. Cells Tissues Organs. 2011;193(4):215-38. doi: 10.1159/000320026.
Gasser RF. The development of the facial nerve in man. Ann Otol Rhinol Laryngol. 1967;76(1):37-56. doi: 10.1177/000348946707600103.
Lobko PI, Khil’kevich SI. Promezhutochnyi nerv i ego mesto v sisteme cherepnykh nervov [The intermediate nerve and its place in the system of cranial nerves]. Arkh Anat Gistol Embriol. 1989;97(9):37-46. Russian.
Marshall H, Nonchev S, Sham MH, Muchamore I, Lumsden A, Krumlauf R. Retinoic acid alters hindbrain Hox code and induces transformation of rhombomeres 2/3 into a 4/5 identity. Nature. 1992;360(6406):737-741. doi: 10.1038/360737a0.
Méndez-Maldonado K, Vega-López GA, Aybar MJ, Velasco I. Neurogenesis from neural crest cells: molecular mechanisms in the formation of cranial nerves and ganglia. Front Cell Dev Biol. 2020;8:635. doi: 10.3389/fcell.2020.00635.
Babuci A, Catereniuc I, Zorina Z, Bendelic A, Botnari T, Stepco E, Lehtman S, Strisca S, Nastas L, Motelica G, Procopenco O. Morphology and variability of the facial nerve trunk depending on the branching pattern, gender, anthropometric type and side of the head in Moldovan population. Folia Morphol (Warsz). 2023;82(4):791-797. doi: 10.5603/FM.a2022.0088.
Cvetko E. Sternocleidomastoid muscle additionally innervated by the facial nerve: case report and review of the literature. Anat Sci Int. 2015;90(1):54-6. doi: 10.1007/s12565-013-0224-8.